

You remember the name of your first-grade teacher. You remember a song lyric from 2009 that you haven't heard in years. But you cannot recall what you ate for lunch last Tuesday, or the face of the person who held the door for you this morning. This asymmetry feels like a malfunction, as though your brain's filing system has gaps where important information should be. For most of the history of neuroscience, researchers treated forgetting exactly this way: as passive decay, a failure of the machinery to hold onto what it once stored.
That framing is wrong. Over the past two decades, a series of discoveries across molecular biology, cognitive psychology, and computational neuroscience have converged on a radically different conclusion: forgetting is not the absence of remembering. It is something your brain does on purpose. The research trail runs from fruit flies in a Florida laboratory to brain-imaging studies at Cambridge to computational models that explain why perfect memory would be a curse, not a gift. Together, these findings tell a story about memory that is far stranger than simple storage and retrieval.
The Forgetting Cells
The first major breakthrough came from an unexpected place: the fruit fly. In 2012, neuroscientist Ronald Davis at the Scripps Research Institute and his longtime collaborator Yi Zhong identified a specific population of dopamine neurons in Drosophila that actively erased memories after they formed. These weren't neurons involved in learning or recall. They were forgetting cells, dedicated circuits whose sole function was to degrade memory traces over time.
The mechanism Davis and Zhong uncovered was remarkably specific. After a fly formed an associative memory (learning to connect an odor with a mild shock, for instance), dopamine neurons continuously signaled to the mushroom body, the fly's memory center, through a receptor called DAMB. This signal activated a protein called Rac1, which in turn triggered cofilin, an enzyme that dismantles the actin cytoskeleton holding synaptic connections in place. The memory trace didn't fade passively. It was actively disassembled, molecule by molecule, by a dedicated biochemical cascade.
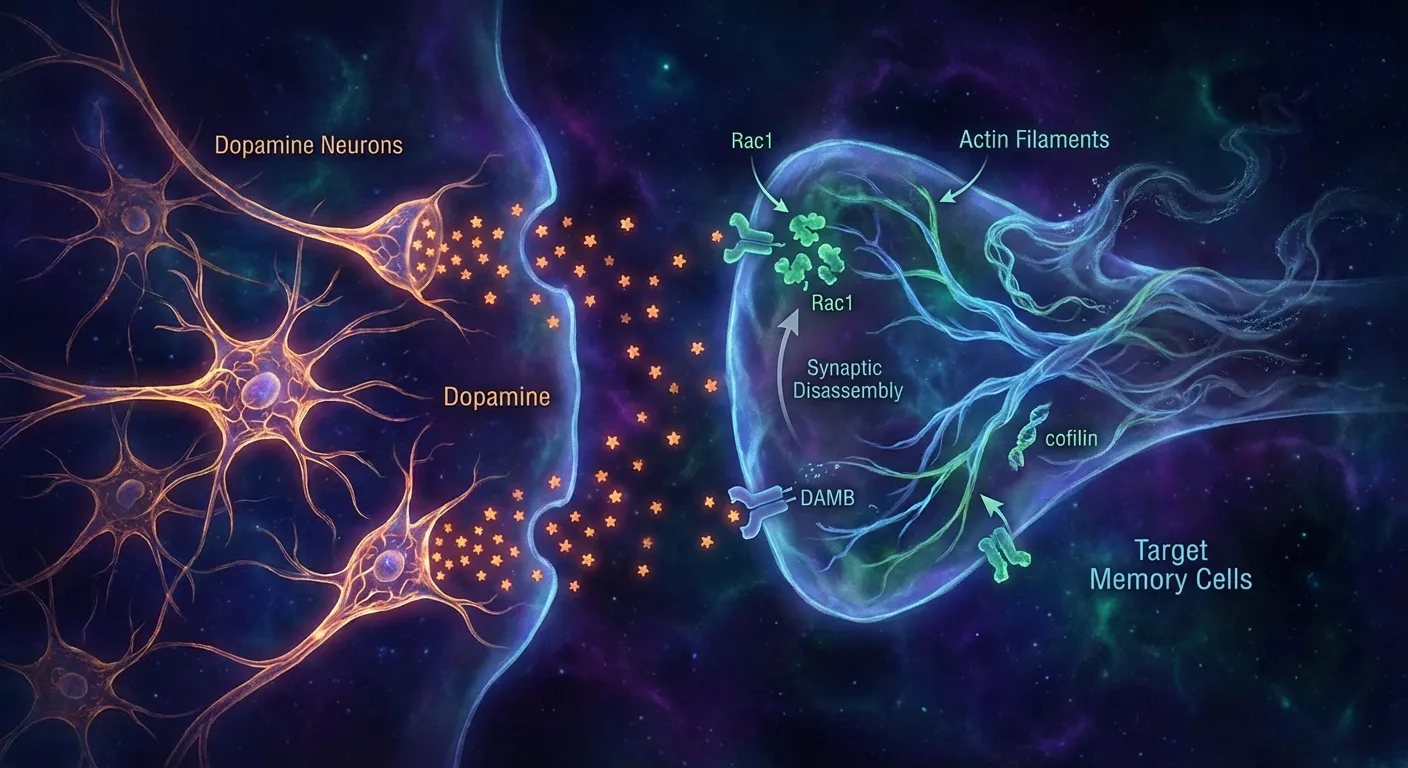
When Davis's team blocked these dopamine signals, flies retained memories far longer than normal. When they amplified the signals, memories vanished faster. The implication was clear: the brain doesn't just build memories and hope they stick. It simultaneously runs an erasure program that controls how long each trace persists. Memory duration isn't determined solely by how strongly something was encoded. It's determined by the balance between the forces of retention and the forces of active forgetting.
Subsequent research confirmed that the same Rac1-dependent forgetting pathway operates in mammals. Studies in mice showed that manipulating Rac1 activity in the hippocampus could extend or shorten memory retention, suggesting that the basic architecture of active forgetting is conserved across hundreds of millions of years of evolution. As Davis wrote in a 2017 review, "The biology of forgetting is not simply the biology of memory in reverse."
Suppression: The Voluntary Erase Button
While Davis and Zhong were mapping the molecular machinery of intrinsic forgetting (the automatic, ongoing process), cognitive psychologist Michael Anderson at the University of Cambridge was discovering something equally striking at the behavioral level. Anderson's work focused on a different kind of forgetting: the deliberate, voluntary suppression of unwanted memories.
Anderson's experimental paradigm, called "think/no-think," works like this. Participants learn pairs of words (for example, "roach" and "ordeal"). Then, during a test phase, they see one word and are either instructed to recall its partner or to actively suppress the partner, pushing it out of consciousness. Afterward, when tested on all the pairs, participants show significantly worse recall for the suppressed items, even when given entirely new cues to retrieve them.
This is not simply a matter of not practicing the memory. The suppressed memories become genuinely less accessible, as though the act of pushing them away has weakened the underlying trace. Anderson's brain imaging studies revealed that during successful suppression, the prefrontal cortex sends inhibitory signals to the hippocampus, damping down activity in exactly the regions responsible for storing the memory. The prefrontal cortex effectively tells the memory system: delete this.
The finding challenged a deeply held assumption in psychology, that memories, once formed, persist indefinitely somewhere in the brain and that forgetting reflects only a failure to retrieve them. Anderson's work suggested instead that retrieval suppression actively degrades the stored representation itself. You can, in a meaningful neurobiological sense, choose to forget.
Reconsolidation: Every Recall Is an Edit
Perhaps the most counterintuitive discovery in the science of memory came from Karim Nader, then a postdoctoral researcher at New York University. In 2000, Nader published a study that upended the textbook understanding of how memories are stored. The prevailing model held that memories undergo a one-time consolidation process, converting from fragile short-term traces into stable long-term ones. Once consolidated, a memory was considered fixed, like data written to a hard drive.
Nader showed this was wrong. When rats recalled a fear memory, the memory became temporarily unstable, requiring a fresh round of protein synthesis to be re-stored. If Nader blocked protein synthesis during this window (using the drug anisomycin injected into the amygdala), the memory was dramatically weakened or erased entirely. The act of remembering didn't just access the memory. It opened it up for editing.
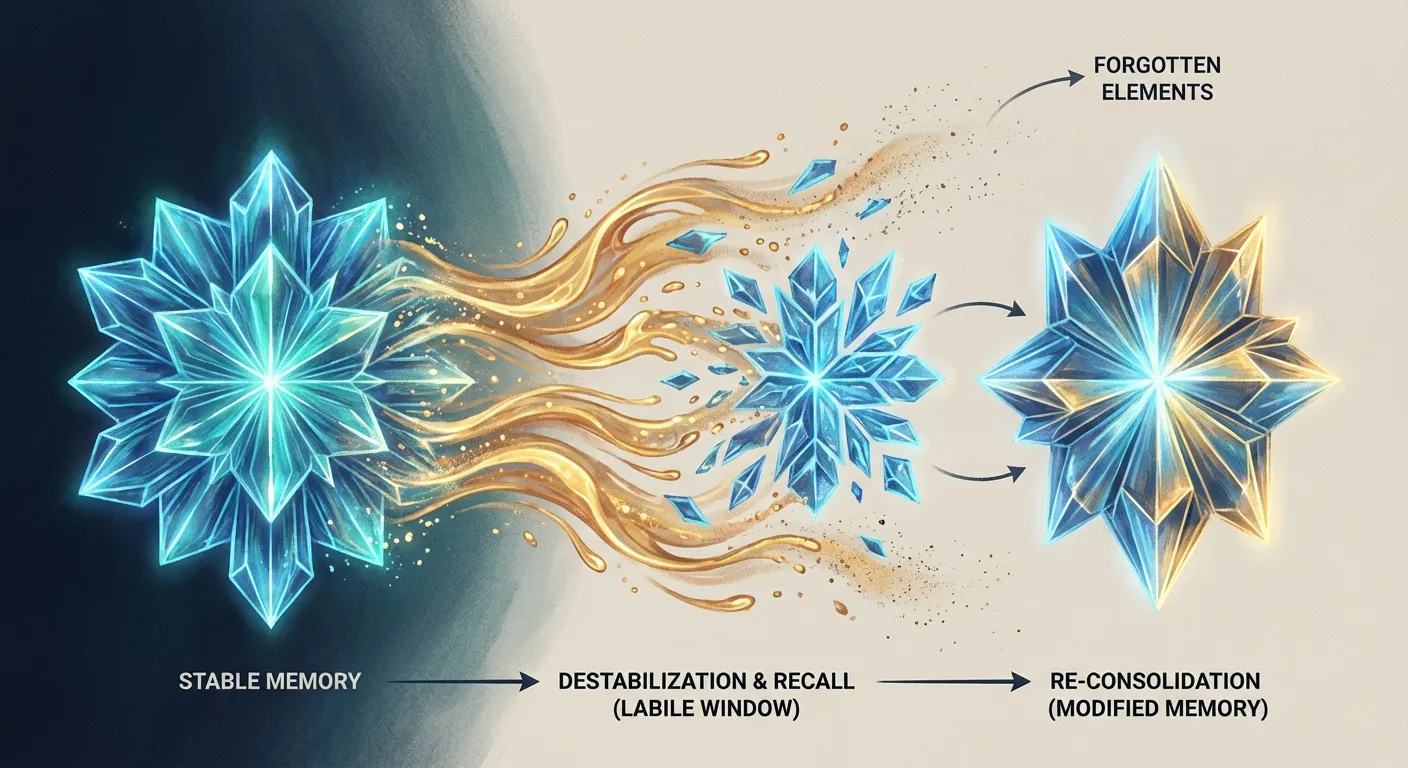
This process, called reconsolidation, means that memories are never truly permanent in the way we once assumed. Every act of recall is an opportunity for the brain to update, modify, or diminish a memory. The relationship between sleep and memory consolidation adds another layer: the brain appears to use sleep to selectively strengthen some memories while allowing others to weaken, a nightly triage process that shapes what you carry forward.
Reconsolidation explains phenomena that were previously mysterious. Therapeutic approaches like exposure therapy work partly because recalling a traumatic memory in a safe context allows the brain to reconsolidate it with a less intense emotional signature. The memory of the event persists, but its visceral power diminishes. This isn't merely psychological habituation. It's a molecular process in which the original trace is literally rewritten.
Why Forgetting Is a Feature
The evidence from Davis's flies, Anderson's suppression experiments, and Nader's reconsolidation work all point in the same direction: the brain has multiple, overlapping systems for weakening or removing memories. Evolution does not invest in redundant systems without reason. The natural question is: why? What advantage does deliberate memory erasure provide?
The most compelling answer comes from computational theory. In the 1990s, researchers studying artificial neural networks discovered that networks trained to remember everything performed poorly on new tasks. They suffered from what computer scientists call overfitting: the network became so specialized to past examples that it couldn't generalize to novel situations. The solution, developed independently by several groups, was dropout, a technique that randomly deactivates a fraction of neural connections during training, forcing the network to develop more robust, generalizable representations.
The parallel to biological forgetting is striking. A brain that retained every sensory detail, every face glimpsed in a crowd, every ambient noise from every room, would be overwhelmed with irrelevant data. Jorge Luis Borges explored this thought experiment in his short story "Funes the Memorious," about a man cursed with perfect memory who finds himself incapable of abstract thought, because thinking requires the ability to forget particulars and extract patterns.
Active forgetting provides exactly this capacity. By continuously pruning low-value memories, the brain maintains the flexibility to learn new information, to generalize from past experience, and to adapt to changing circumstances. The ways AI systems handle information overload mirror this biological strategy: both artificial and biological systems must balance retention against adaptability, and both solve the problem by deliberately discarding information.
Clinical Implications: When Forgetting Fails
Understanding active forgetting as a feature rather than a bug reframes several psychiatric conditions. Post-traumatic stress disorder, for instance, can be understood partly as a failure of the brain's forgetting machinery. Traumatic memories that should be gradually weakened by intrinsic forgetting instead persist with full emotional intensity, intruding on consciousness unbidden. If the dopamine-Rac1 pathway that Davis identified in flies operates similarly in humans (and accumulating evidence suggests it does), then PTSD may involve a specific breakdown in the molecular cascade responsible for memory erasure.
Addiction presents a related challenge. Drug-associated memories, the cues and contexts linked to substance use, are notoriously resistant to extinction. The reconsolidation window offers a potential therapeutic target: if drug memories could be reactivated and then disrupted during their vulnerable phase, the cravings they trigger might be diminished. Early clinical trials using propranolol (a beta-blocker that interferes with reconsolidation) to treat both PTSD flashbacks and addiction cravings have shown promising, though still preliminary, results.
Conversely, conditions involving excessive forgetting, like Alzheimer's disease, may involve a pathological overactivation of forgetting pathways rather than simply a failure to form new memories. Research published in Molecular Psychiatry in 2024 found connections between active forgetting mechanisms and several neuropsychiatric diseases, suggesting that the balance between remembering and forgetting may be disrupted in fundamentally different ways across different conditions.
Where This Leads
What makes these findings collectively powerful is that they operate at different scales and through different mechanisms, yet all serve the same function. Dopamine-driven intrinsic forgetting works automatically in the background. Retrieval suppression is a voluntary, top-down process. Reconsolidation edits memories at the moment of recall. The brain does not rely on a single eraser. It has an entire toolkit, each instrument suited to a different kind of memory management.
This understanding opens practical possibilities that would have seemed like science fiction a generation ago. Targeted forgetting therapies could one day help people with PTSD shed the emotional charge of traumatic memories without erasing the memories themselves. Reconsolidation-based interventions could reshape addictive associations. Even educational strategies might be redesigned around the insight that some forgetting is necessary for deep learning, that the student who forgets certain details may actually understand the material better than the student who remembers everything.
The deepest implication, though, may be philosophical. We tend to think of our identities as the sum of what we remember. But if the brain is actively curating that collection, continuously deciding which memories survive and which are dissolved, then identity is as much about what we forget as what we retain. The self is not a complete archive. It is an edited story, shaped as much by deletion as by addition, and the editor is a set of molecular pathways that have been refining their craft for hundreds of millions of years.
Sources
- The Biology of Forgetting: A Perspective (Davis, 2017)
- To Remember, the Brain Must Actively Forget (Quanta Magazine)
- Neural, Cellular and Molecular Mechanisms of Active Forgetting (Bhatt et al., 2018)
- Active Forgetting and Neuropsychiatric Diseases (Molecular Psychiatry, 2024)
- Reconsolidation and the Dynamic Nature of Memory (Nader & Hardt, 2009)